Inflammation is simply a physiologic response process
generated by the body in response to injury, infection, or
irritation. In acute stages, the inflammatory process is
vital to the healing process; however, chronic inflammation
can increase disease- associated morbidity. New insights
into the chronic inflammatory process now provide evidence
that this mechanism is a negative contributor to an
ever-expanding list of chronic conditions, including
Alzheimer's disease, cardiovascular diseases, diabetes,
asthma, cancer, and even depression.
Epidemiological studies have shown that chronic
inflammation predisposes individuals to various types of
cancer and the host response to malignant disease shows
several parallels with inflammation and wound healing. It is
estimated that underlying infections and inflammatory
responses and linked to 15-30% of all death from cancer
worldwide. The functional relationship between
inflammation and cancer is not new. In 1863, Virchow
hypothesized that the origin of cancer was at sites of
chronic inflammation, in part based on his hypothesis
that some classes of irritants, together with the tissue
injury and ensuing inflammation they cause, enhance cell
proliferation (1). Today, the causal relationship
between inflammation, innate immunity and cancer is more
widely accepted.
In a sense, tumours act as wounds that fail to heal (2).The hallmarks of cancer-related inflammation include the
presence of inflammatory cells and inflammatory mediators
(for example, chemokines, cytokines and prostaglandins) in
tumour tissues, tissue remodelling and angiogenesis (growth
of new blood vessels) similar
to that seen in chronic inflammatory responses, and tissue
repair.
Cytokines A small protein released by
cells that has a specific effect on the interactions
between cells, on communications between cells or on the
behavior of cells. The cytokines includes the
interleukins, lymphokines and cell signal molecules,
such as tumor necrosis factor and the interferons, which
trigger inflammation and respond to infections.
Chemokine: One of a large group
of proteins that act as lures and were first found
attracting white blood cells. The chemokines are involved in a
wide variety of processes including acute and chronic types of
inflammation, infectious diseases, and cancer.
Chemokines may lure cancer cells and help determine the
sites to which cancer cells spread by metastasis.
Prostaglandin: One of a number
of hormone-like substances that participate in a wide
range of body functions such as the contraction and
relaxation of smooth muscle, the dilation and
constriction of blood vessels, control of blood
pressure, and modulation of inflammation. Prostaglandins
are derived from a chemical called arachidonic acid.
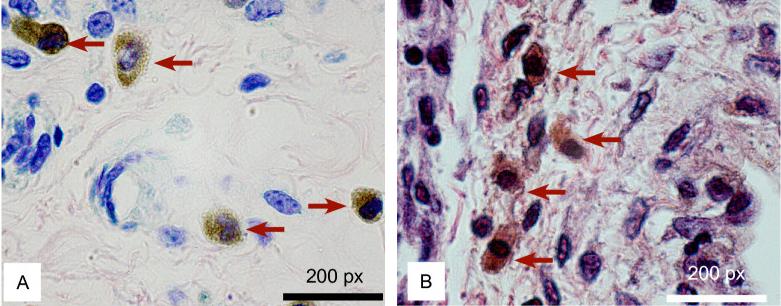
Presence
of LYVE-1-positive macrophages during bladder cancer
progression. Representative photomicrographs
indicating LYVE-1-positive cells. Tumour associated
macrophages (TAMS) are indicated by a red arrow.
Evidence currently available suggests that in established,
progressively growing solid tumours, tumour associated
macrophages (TAMs) are reprogrammed to induce immune
suppression of host defenses in situ, through
release of specific cytokines, prostanoids and other
humoral mediators. This disordered response, results in
the inhibition of effective anti-cancer cell-mediated
immune mechanisms. Concurrently, TAMs produce tumour
growth promoting factors. The summation of this complex
interplay of biological factors results in progressive
tumour growth and tumour cell dissemination.
An overview of inflammation and carcinogenesis
A key attribute of an effective Immune system is to be
able to detect the presence of trouble - this could be dead
or damaged cells, tumour cells or infection with viruses,
bacteria or eukaryotic parasites. Some aspects of this have
already been dealt with; acute inflammation is the major
system for sensing trauma. However, the Immune system needs
to be able to detect more subtle changes and also needs to
encourage a two-way communication between the innate and
adaptive immune responses.
In terms of protection against infection the ability of
certain cells to detect the presence of prokaryotic
molecules (like bacteria) is of primary importance. Though
there is considerable sophistication and subtlety to the
mechanisms which do this we can focus here on 3 cells types:
mast, macrophages and dendritic cells.
Typical primary immune system response to an assault
Mast cells primarily detect danger via receptors for
activated complement. They also use antibodies as sensing
tools via Fc receptors for both IgE (high affinity) and IgG
(low affinity).
The two cell types I want to emphasise here are the
Macrophage and Dendritic cell. These cells share a
number of receptors which bind structures which are
specific to bacteria or fungi, mostly carbohydrates. In
fact blood monocytes may be induced to differentiate
into either macrophages or dendritic cells under
appropriate distinct conditions. The response of the
cells to the ligation of these receptors includes
phagocytosis and other type of responses but the
critical issues relevant in this context are
cytokine production
antigen presentation
Macrophages are exquisitely sensitive to the
lipopolysaccharides (LPS) produced by certain bacteria. They
respond by producing cytokines notably TNFalpha, but also
IL1 and IL6. These mediators induce the Acute Phase
Response, which is a rapid systemic response which massively
increases the concentration of many key serum proteins to
aid the host defence response. C reactive protein (CRP) and
Mannose binding protein are natural activators of the
Complement system.
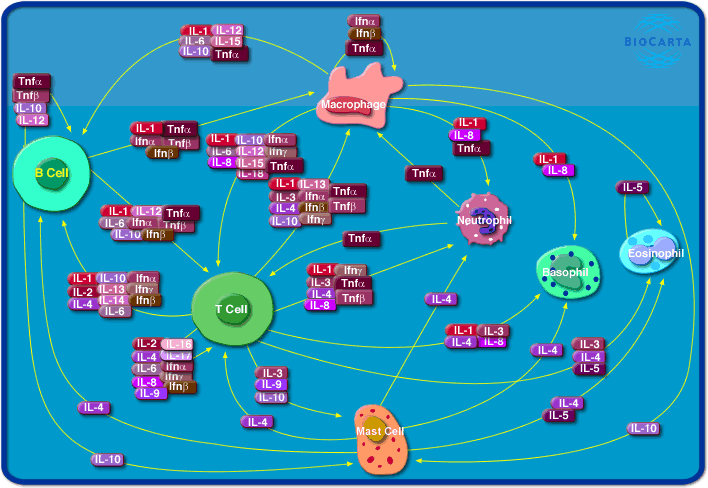
The previous image shows a summary of the
important cells and molecules in the human immune system -
the top half of the picture represents detection of invaders, and the
bottom half represents the defence which is triggered
by that detection.
The immune system cell communication network
Several different cell types coordinate their efforts as
part of the immune system, including B cells, T cells,
macrophages, neutrophils, basophils and eosinophils. Each of
these cell types has a distinct role in the immune system,
and communicates with other immune cells using secreted
factors called cytokines, including interleukins, TNF, and
the interferons. Macrophages phagocytose foreign bodies and
are antigen-presenting cells, using cytokines to stimulate
specific antigen dependent responses by B and T cells and
non-specific responses by other cell types. T cells secrete
a variety of factors to coordinate and stimulate immune
responses to specific antigen, such as the role of helper T
cells in B cell activation in response to antigen. The
proliferation and activation of eosinophils, neutrophils and
basophils respond to cytokines as well. Cytokine
communication is often local, within a tissue or between
cells in close proximity. Each of the cytokines is secreted
by one set of cells and provokes a response in another
target set of cells, often including the cell that secretes
the cytokine.
Some cytokines, like IL-1, interferons and TNF, stimulate a
broad inflammatory response in response to infection or
injury. Other cytokines have more specific functions such
the following examples. IL-2 stimulates the proliferation
and activation of B and T cells. IL-4 plays a role in the
differentiation of Th2 cells, in allergic responses, and in
the switching of antibody types. IL-5 stimulates the
production and maturation of eosinophils during
inflammation. IL-8 is a chemokine, a chemotactic factor that
attracts neutrophils, basophils and T cells to sites of
inflammation. IL-12 and IL-18 are involved in helper T cell
differentiation. IL-10 apparently acts to repress secretion
of proinflammatory cytokines. The complex interplay of these
different cytokine functions with immune cells is essential
for correct immune function.
Role of inflammation in the evolution of cancer
To understand the role of inflammation in the evolution of cancer, it is
important to understand what inflammation is and how it contributes to physiological and pathological
processes such as wound healing and infection (Fig. 1). In response to tissue injury, a
multifactorial network of chemical signals initiate and maintain a host response designed to 'heal' the
afflicted tissue. This involves activation and directed migration of leukocytes (neutrophils,
monocytes and eosinophils) from the venous system to sites of damage, and tissue mast cells also have a
significant role.
When tissue homeostasis is perturbed, sentinel
macrophages and mast cells immediately release soluble
mediators, such as cytokines, chemokines matrix remodelling
proteases and reactive oxygen species (ROS), and bioactive
mediators such as histamine, that induce mobilization and
infiltration of additional keucocytes into damaged tissue (a
process that is known as inflammation). Macrophages and mast
cells can also activate vascular and fibroblast responses in
order to orchestrate the elimination of invading organisms
and initiate local tissue repair.
Acute activation of innate immunity sets the stage for
activation of the more sophisticated adaptive immune system.
Induction of efficient primary adaptive immune responses
requires direct interactions with mature antigen-presenting
cells and pro-inflammatory milieu.
Together, acute activation of these distinct
immune-response pathways efficiently removes or eliminates
invading pathogens, damaged cells and extracellular matrix
(ECM). In addition, once assaulting agents are eliminated,
immune cells are crucially involved in normalizing
cell-proliferation and cell death pathways to enable re-epitheliarization
and new extracellular matrix synthesis. Once wound healing
is complete, inflammation resolves and tissue homeostasis
returns.
In (a) a normal skin
tissue have a highly organized and segregated architecture.
Epithelial cells sit atop a basement membrane separated from
the vascularized stromal (dermis) compartment. Upon wounding
or tissue assault, platelets are activated and release
vasoactive mediators that regulate vascular permeability,
influx of serum fibrinogen, and formation of the fibrin
clot. They also release proteolytic enzymes necessary for
remodelling of extracellular matrix. In combination,
granulocytes, monocytes and fibroblasts are recruited, the
venous network restored, and re-epithelialization across the
wound occurs. Epithelial and stromal cell types engage in a
reciprocal signalling dialogue to facilitate healing. Once
the wound is healed, the reciprocal signalling subsides.
In (b) an invasive carcinomas is less
organized. Neoplasia-associated angiogenesis and
lymphangiogenesis produces a chaotic vascular organization
of blood vessels and lymphatics where neoplastic cells
interact with other cell types (mesenchymal, haematopoietic
and lymphoid) and a remodelled extracellular matrix.
Neoplastic cells produce an array of cytokines and
chemokines that are mitogenic and/or chemoattractants for
granulocytes, mast cells, monocytes/macrophages, fibroblasts
and endothelial cells. In addition, activated fibroblasts
and infiltrating inflammatory cells secrete proteolytic
enzymes, cytokines and chemokines, which are mitogenic for
neoplastic cells, as well as endothelial cells involved in
neoangiogenesis and lymphangiogenesis. These factors
potentiate tumour growth, stimulate angiogenesis, induce
fibroblast migration and maturation, and enable metastatic
spread via engagement with either the venous or lymphatic
networks.
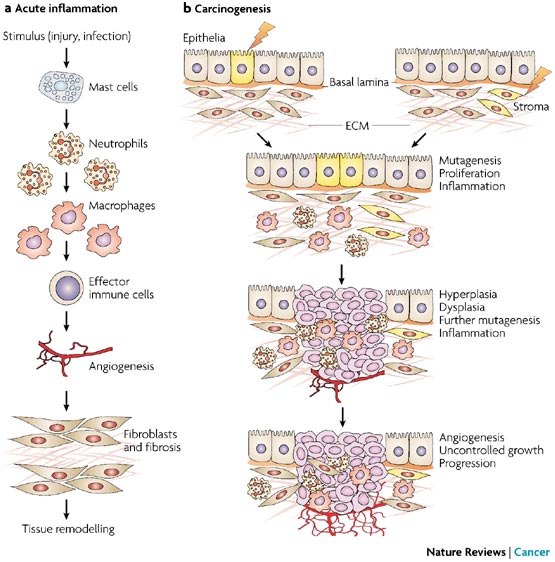
a | The sequence of events in
acute inflammation and tissue repair. The process of
inflammation initiates a series of catabolic and anabolic
processes that occur in a defined order, first eliminating
foreign pathogens and then remodelling tissue, thereby
establishing homeostasis. Shown in this figure are: first,
the activation of resident cells (mast cells, resident
macrophages and dendritic cells) and rapid entry of
granulocytes in response to injury; second, further
recruitment of macrophages; third, infiltration of effector
immune cells (lymphocytes) to enable specific immune
responses; fourth, the recruitment and activation of
mesenchymal cells such as endothelial cells and fibroblasts
to form new blood vessels and a collagenous matrix; and
fifth, tissue remodelling. In its initial stages,
inflammation is an aggressive state that can destroy both
exogenous pathogens and host tissues; this is followed by a
switch to a state that promotes cell survival and tissue
regeneration. b | Carcinogenesis as
the chaotic disorganization of inflammation and repair. In
contrast to the orderly series of events shown in part
a, during chronic unresolved
inflammation and carcinogenesis these events are chaotically
disorganized and homeostasis is not achieved. During
carcinogenesis, both epithelial and stromal elements might
initially undergo alterations that promote epithelial cell
proliferation and mutation. This alteration in tissue
homeostasis can in turn lead to an inflammatory response,
which then further promotes tumour growth through the
activation of the surrounding stroma, especially
neovascularization. Continued hyperplasia and dysplasia
eventually lead to an invasive neoplastic state. The process
shown here has been described with the metaphor that
"tumours are wounds that do not heal". Both epithelial cells
and cells of the microenvironment are targets for
chemopreventive agents at all the steps shown.
Chemopreventive agents might be anti-mutagenic, anti-proliferative,
anti-inflammatory or anti-angiogenic, therefore restoring
tissue homeostasis that has been disrupted during
carcinogenesis. Early genetic or epigenetic changes in
epithelia or stroma are shown in yellow. ECM, extracellular
matrix.
Two arms of the immune system - the innate and the
adaptive - are exquisitely well adapted for fighting
pathogens, but their role in combating cancer is
decidedly more paradoxical. The innate system furnishes
an initial inflammatory response to a microbial insult
by attacking any invading pathogen indiscriminately,
whereas adaptive immunity furnishes a delayed response
that homes in on a particular pathogen. In cancer, both
systems may sometimes attack tumour cells. But a tumour
protects itself by recruiting the innate system to
enhance its development.
In the late 1990s, Frances Balkwill of the institute of
Cancer at Queen Mary, University of London, had been doing
research on a cytokine known as tumour necrosis factor
(TNF), which was named for its ability to kill cancer cells
when administered directly into a tumour at high levels. But
when TNF lingers as a chronic, low level presence in the
tumour, it acts very differently. Balkwill's lab turned off
the TNF gene in mice so that the rodents could not produce
the protein: to their surprise, the mice did not contract
tumours. All the people who were working on TNF as an
anticancer agent were horrified. This cytokine they thought
was a treatment for cancer was actually working as an
endogenous tumour promoter.
The ready availability of knockout mice, in which the
effects of selectively switching off genes could be tested,
helped to highlight the cancer-inflammation link. Coussens
and her UCSF colleagues Douglas Hanahan and Zena Werb
reported in 1999 that mice engineered with activated cancer
genes but without mast cells developed premalignant tissue
that did not progress to full malignancy. In 2001, Jeffrey
W. Pollard and his co-workers at the Albert Einstein College
of Medicine described mice that were genetically engineered
to be susceptible to breast cancer tumours but that produced
precancerous tussue that did not turn malignant unless it
enlisted the assistance of macrophages.
Tumour cells produce various cytokines and chemokines
that attract leukocytes. The inflammatory component of a
developing neoplasm may include a diverse leukocyte
population - for example, neutrophils, dendritic cells,
macrophages, eosinophils and mast cells, as well as
lymphocytes - all of which are capable of producing an
assorted array of cytokines, cytotoxic mediators including
reactive oxygen species, serine and cysteine proteases, MMPs
and membrane-perforating agents, and soluble mediators of
cell killing, such as TNF-,
interleukins and interferons (7) (8).
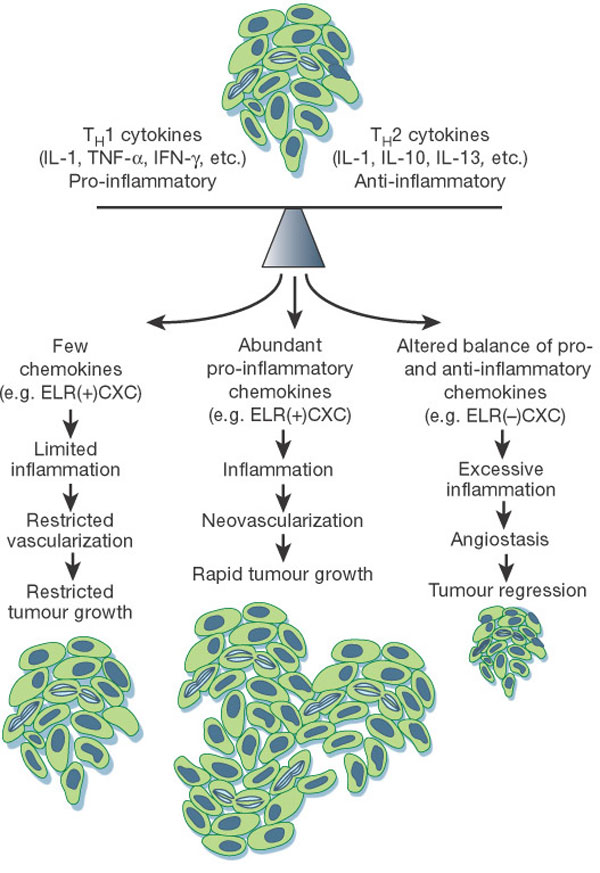
The balance of cytokines in any given tumour is critical
for regulating the type and extent of inflammatory
infiltrate that forms. Tumours that produce little or no
cytokines or an overabundance of anti-inflammatory cytokines
induce limited inflammatory and vascular responses,
resulting in constrained tumour growth. In contrast,
production of an abundance of pro-inflammatory cytokines can
lead to a level of inflammation that potentiates
angiogenesis, thus favouring neoplastic growth.
Alternatively, high levels of monocytes and/or neutrophil
infiltration, in response to an altered balance of pro-
versus anti-inflammatory cytokines, can be associated with
cytotoxicity, angiostasis and tumour regression. In tumours,
interleukin-10 is generally a product of tumour cells and
tumour-associated macrophages.
Diet and the impact on inflammation and cancer
Writing note: include here
some introduction text. I have to bridge to omega 3 and
other immunomodulators. The key point is to demonstrate the
immunotherapeutic factors of these foods/supplements. If I
have enough time I should add the work performed by Dr
Nobuto Yamamoto and how to turn TAMS into cancer cell
enemies (if I finally get an interview with him or his son).
Could beta glucan do the same? An hypothesis to explore. I
should also write about cytokines, stress and immunity. I
probably should include more explanation about the other
facet of this topic, the eicosanoids especially because of
their importance in the context of omega 3.
Omega 3 as a modulator of inflammation
Among the fatty acids, it is the omega-3 polyunsaturated
fatty acids (PUFA) which possess the most potent
immunomodulatory activities, and among the
omega-3 PUFA, those from fish oil—eicosapentaenoic
acid (EPA) and docosahexaenoic acid (DHA)—are more
biologically potent than
-linolenic
acid (ALA).
Some of the effects of omega-3 PUFA
are brought by modulating of the amount and types
of eicosanoids made, and other effects are elicited by
eicosanoid-independent mechanisms, including
actions upon intracellular signaling pathways,
transcription factor activity and gene expression. Animal
experiments and clinical intervention studies
indicate that omega-3 fatty acids have
anti-inflammatory properties and, therefore, might
be useful in the management of inflammatory and
autoimmune diseases.
Coronary heart disease,
major depression, aging and cancer are
characterized by an increased level of interleukin 1 (IL-1),
a proinflammatory cytokine. Similarly, arthritis,
Crohn’s disease, ulcerative colitis and lupus
erythematosis are autoimmune diseases
characterized by a high level of IL-1 and the
proinflammatory leukotriene LTB4
produced by omega-6 fatty acids. There have been
a number of clinical trials assessing the benefits of
dietary supplementation with fish oils in several
inflammatory and autoimmune diseases in humans,
including rheumatoid arthritis, Crohn’s disease,
ulcerative colitis, psoriasis, lupus erythematosus,
multiple sclerosis and migraine headaches. Many of the
placebo-controlled trials of fish oil in chronic
inflammatory diseases reveal significant benefit,
including decreased disease activity and a lowered
use of anti-inflammatory drugs.
The brain as an immune system modulator
New
field of research, known as psychoneuroimmunology, is
exploring how the immune system and the brain may interact
to influence health. For years stress has been suspected of
increasing susceptibility to various infectious diseases or
cancer. Now evidence is mounting that the immune system and
the nervous system may be inextricably interconnected.
Research has shown that a wide range of stresses, from
losing a spouse to facing a tough examination, can deplete
immune resources, causing levels of B and T cells to drop,
natural killer cells to become less responsive, and fewer
IgA antibodies to be secreted in the saliva.
Biological links between the immune system and the
central nervous system exist at several levels. One
well-known pathway involves the adrenal glands, which, in
response to stress messages from the brain, release
corticosteroid hormones into the blood. In addition to
helping a person respond to emergencies by mobilizing the
body's energy reserves, these "stress hormones" decrease
antibodies and reduce lymphocytes in both number and
strength.
More recently, it has become apparent that hormones and
neuropeptides (hormone-like chemicals released by nerve
cells), which convey messages to other cells of the nervous
system and organs throughout the body, also "speak" to cells
of the immune system. Macrophages and T cells carry
receptors for certain neuropeptides; natural killer cells,
too, respond to them. Even more surprising, some macrophages
and activated lymphocytes actually manufacture typical
neuropeptides. At the same time, some lymphokines, secreted
by activated lymphocytes such as interferon and the
interleukins, can transmit information to the nervous
system. Hormones produced by the thymus, too, act on cells
in the brain.
In addition, the brain may directly influence the immune
system by sending messages down nerve cells. Networks of
nerve fibers have been found that connect to the thymus
gland, spleen, lymph nodes, and bone marrow. Moreover,
experiments show that immune function can be altered by
actions that destroy specific brain areas.
Audio Visual Documentation
These two following videos, even tinted with some
pharmaceutical references are very interesting presentation
illustrating the relationship between the immune system and
cancer.
Lisa
Coussens is Professor of Pathology at the University of
California, San Francisco (UCSF) and Co-Director of the
Mouse Pathology Core Facility at the UCSF Helen Diller
Family Comprehensive Cancer Center. Her training included
seven years at Genentech Inc in the 1980s, where she
participated in the cloning and characterization of receptor
tyrosine kinases; PhD training in biological chemistry at
the University of California, Los Angeles, USA; and
postdoctoral work in Douglas Hanahan's laboratory at UCSF.
Her research focuses on the role of inflammatory cells
and leukocyte proteases as critical regulators of skin, lung
and breast cancer development. By studying mouse models of
cancer development, the Coussens lab is identifying crucial
molecules that are involved in regulating tumour-associated
inflammation, matrix remodelling and angiogenesis.
Identification of these important regulatory molecules
reveals drug targets that can be used to design novel
therapeutic strategies for arresting cancer development in
humans.
Lisa Coussens is recipient of numerous awards, including
the V Foundation Scholar award (2000), the Malinckrodt Award
for Medical Research (2000), the AACR Gertrude B Elion
Cancer Research Award (2002), and the Era of Hope Scholar
Award (2006).
Dr.
Michael Karin is currently a Professor of Pharmacology at
the School of Medicine at the University of California, San
Diego. He received his Ph.D. in Molecular Biology from UCLA
in 1979. He is a leading world authority on signal
transduction pathways that regulate gene expression in
response to extracellular stimuli. Key achievements include
definition of cis elements that mediate gene induction by
hormones, cytokines and stress, identification and
characterization of the transcription factors that recognize
these elements and the protein kinase cascades that regulate
their activities. He has published over 200 scientific
articles and is an inventor on over 14 different patents or
pending patent applications. Recently Dr. Karin was ranked
first worldwide by the Institute of Scientific Information
(ISI) in a recent listing of most-cited molecular biology
and genetic research papers published in prestigious
journals.
References
(1) Balkwill F.& Mantovani A. Inflammation and cancer:
back to Virchow? Lancet 357, 539-545 (2001)
(2) Dvorak, H. F.
Tumors: wounds that do not heal. Similarities between tumor
stroma generation and wound healing. N. Engl. J. Med.315, 1650-1659 (1986).
(3) Karin E. de Visser et al. Paradoxical
roles of the immune system during cancer development. Nature
Cancer review vol 6 January 2006 24-37
(4) Alberto Mantovani. Cancer-related
inflammation. Nature Vol 454 July 2008 436-444
(5) Lisa Coussens & Zena Werb. Inflammation
and cancer. Nature vol. 420 December 2002 860-273
(6) Seth Rakoff-Nahoum. Why Cancer and
Inflammation? Yale journal of biology and medicine 79 (2006)
123-130
(7) Kuper H. et al. Infections as a major
preventable cause of human cancer. j. Intern. Med. 248,
171-183 (2000)
(8) Wahl L. et al. Tumor associated
macrophages as targets for cancer therapy. J. Natl Cancer
Inst. 90, 1583-1584 (1998)








 -linolenic
acid (ALA).
-linolenic
acid (ALA).  New
field of research, known as psychoneuroimmunology, is
exploring how the immune system and the brain may interact
to influence health. For years stress has been suspected of
increasing susceptibility to various infectious diseases or
cancer. Now evidence is mounting that the immune system and
the nervous system may be inextricably interconnected.
New
field of research, known as psychoneuroimmunology, is
exploring how the immune system and the brain may interact
to influence health. For years stress has been suspected of
increasing susceptibility to various infectious diseases or
cancer. Now evidence is mounting that the immune system and
the nervous system may be inextricably interconnected.  Lisa
Coussens is Professor of Pathology at the University of
California, San Francisco (UCSF) and Co-Director of the
Mouse Pathology Core Facility at the UCSF Helen Diller
Family Comprehensive Cancer Center. Her training included
seven years at Genentech Inc in the 1980s, where she
participated in the cloning and characterization of receptor
tyrosine kinases; PhD training in biological chemistry at
the University of California, Los Angeles, USA; and
postdoctoral work in Douglas Hanahan's laboratory at UCSF.
Lisa
Coussens is Professor of Pathology at the University of
California, San Francisco (UCSF) and Co-Director of the
Mouse Pathology Core Facility at the UCSF Helen Diller
Family Comprehensive Cancer Center. Her training included
seven years at Genentech Inc in the 1980s, where she
participated in the cloning and characterization of receptor
tyrosine kinases; PhD training in biological chemistry at
the University of California, Los Angeles, USA; and
postdoctoral work in Douglas Hanahan's laboratory at UCSF. Dr.
Michael Karin is currently a Professor of Pharmacology at
the School of Medicine at the University of California, San
Diego. He received his Ph.D. in Molecular Biology from UCLA
in 1979. He is a leading world authority on signal
transduction pathways that regulate gene expression in
response to extracellular stimuli. Key achievements include
definition of cis elements that mediate gene induction by
hormones, cytokines and stress, identification and
characterization of the transcription factors that recognize
these elements and the protein kinase cascades that regulate
their activities. He has published over 200 scientific
articles and is an inventor on over 14 different patents or
pending patent applications. Recently Dr. Karin was ranked
first worldwide by the Institute of Scientific Information
(ISI) in a recent listing of most-cited molecular biology
and genetic research papers published in prestigious
journals.
Dr.
Michael Karin is currently a Professor of Pharmacology at
the School of Medicine at the University of California, San
Diego. He received his Ph.D. in Molecular Biology from UCLA
in 1979. He is a leading world authority on signal
transduction pathways that regulate gene expression in
response to extracellular stimuli. Key achievements include
definition of cis elements that mediate gene induction by
hormones, cytokines and stress, identification and
characterization of the transcription factors that recognize
these elements and the protein kinase cascades that regulate
their activities. He has published over 200 scientific
articles and is an inventor on over 14 different patents or
pending patent applications. Recently Dr. Karin was ranked
first worldwide by the Institute of Scientific Information
(ISI) in a recent listing of most-cited molecular biology
and genetic research papers published in prestigious
journals.
